

Images

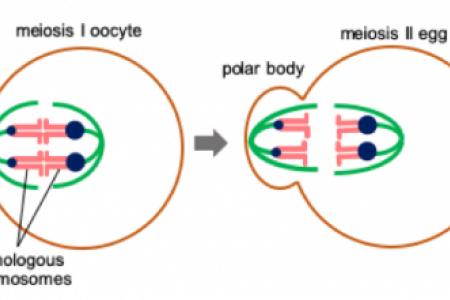
Schematics of meiotic drive in female meiosis.
Schematics of meiotic drive in female meiosis. Asymmetry in female meiosis provides unique opportunity for selfish centromeres to cheat. The spindle is positioned close to the cortex and orients perpendicular to the cortex. Centromeres (blue circles) interact with the spindle and direct chromosome segregation. Selfish centromeres (bigger blue circles) preferentially orient to the interior side of the spindle, which remains in the egg.

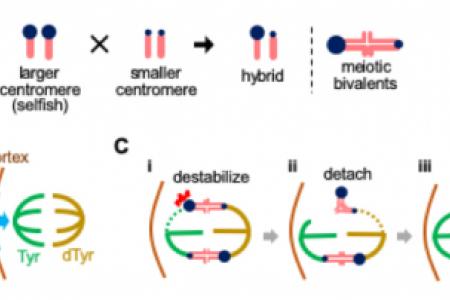
Cell biology of centromere drive.
Cell biology of centromere drive. (A) Crossing different mouse strains generates a hybrid with a difference in centromeres within the bivalent. Larger centromeres with expanded satellite repeats act selfishly over smaller centromeres. (B) Spindle asymmetry depends on active CDC-42 at the polarized cortex. (C) A model for biased orientation of selfish centromeres. Orientation is initially random (i). However, selfish centromeres (bigger blue circles) preferentially destabilize MT attachments with the cortical side (green lines) and detach (ii), leading to biased flipping to the egg side (yellow lines), and biased orientation (iii).

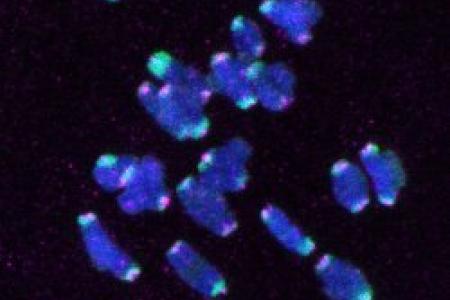
Meiotic bivalents pairing
Meiotic bivalents pairing homologous chromosomes from different species in inter-specific hybrid mouse oocytes

Spindle asymmetry
Spindle asymmetry (grey) induced by polarized cortex (green) serves as a spatial cue to guide selfish centromeres.

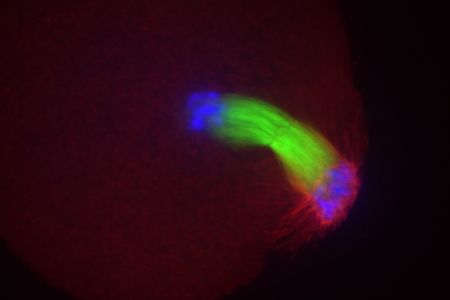
Meiotic drive
Meiotic drive of non-centromeric locus (green)

Centromeres
Selfish centromeres cheating in action!

Mouse oocyte cells
Mouse oocyte cells extruding the polar body, which is an evolutionary dead end for chromosomes.



Meet the Team

Takashi Akera, Ph.D.
Dr. Takashi Akera graduated from the University of Tokyo with a Ph.D. in Biophysics and Biochemistry in 2014. He conducted postdoctoral research at the University of Pennsylvania in the laboratory of Dr. Michael Lampson from 2015 to 2019. He received both Holtzer Award for outstanding postdoctoral research in Cell and Developmental Biology and Kaushal Award for excellence in postdoctoral research in Genetics from the University of Pennsylvania in 2018. He was also a finalist for the ASCB Porter Prize for Research Excellence Award in 2018. Dr. Akera joined the NHLBI in 2019 as an Earl Stadtman tenure-track Investigator and is a member of the American Society for Cell Biology.
Contact the lab

Betsy Clark, PhD

Warif El Yakoubi, PhD

Eddie (Bo) Pan, PhD

Zaak Walton

Takaya Totsuka, Ph.D.

Duílio Silva, PhD

Sebastian Khan
